HLAをより深く知る
HLAとMHC
HLAは発見当初、白血球のみに存在すると考えられてきたが、その後多くの研究がなされ、白血球抗原としてだけでなく、ヒトの主要組織適合性複合体(MHC; Major Histocompatibility Complex)として知られるようになりました(ヒトに関しては、MHC = HLA)。
また、HLAは体のほとんど全ての細胞表面で発現がみられることもわかっています。
HLAの構造
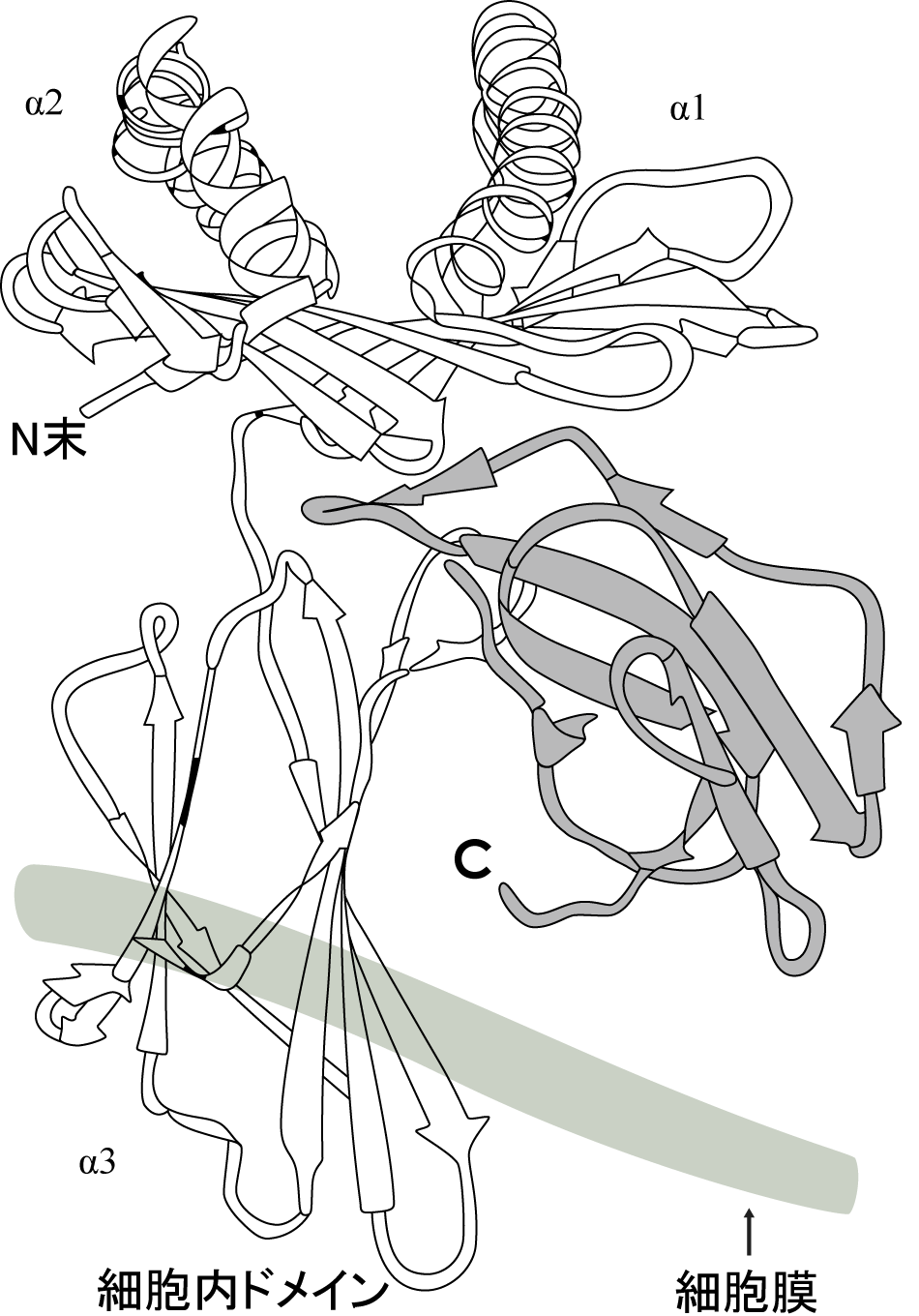 HLA分子はそれぞれの部位でいろいろな機能を分担しています。図をご覧下さい。
HLA分子はそれぞれの部位でいろいろな機能を分担しています。図をご覧下さい。
HLA分子は一部が細胞膜に埋め込まれたようにして、細胞に表現されていますから、細胞外ドメイン、細胞膜貫通ドメイン、細胞内ドメインの3つに分けられ、さらに細胞外ドメインは細胞膜から遠い位置(遠位ドメイン)と膜に近い近位ドメインがそれぞれに異なる機能を持っています。
遠位ドメインはHLA分子の最も重要な機能である、抗原ペプチドの選択的受容と、T細胞への抗原提示をしています。近位ドメインにはT細胞にある分子;CD4・CD8と特異的に結合する構造をもっています。 膜貫通ドメインにはあまり多型性はありません。細胞内ドメインには量的な多型性が知られていて、それは細胞内にあるHLA分子の長短として表現されます。
HLAの多型性は上記のように「表現される」アミノ酸多型性以外に、HLA遺伝子からHLA分子への転写・翻訳を量的に制御する「プロモーター領域」にも多型性が知られています。
ヒトMHC(= HLA)の多型性
白血球抗原型として出発したHLAですが、その多型性は血液型の範疇をはるかに超える特徴をもっています。血液型とは次元の異なる多型として理解すべきでしょう。
血液型は偶然の突然変異がたまたま定着して「型」になったものがほとんどです。そしてその「型」も至極単純で、プラスかマイナスかで分類できるようなものが多く、2または3個の対立遺伝子(アレル)の組み合わせにすぎません。
HLAは「自他認識のマーカー」ですから、非常に多様な「他(た)」を自己から区別するために独特の進化をしました。つまり、ウイルス、細菌などを含むパラサイト(=「他(た)」)は宿主(しゅくしゅ)と共生するために、刻々変異して多様性が蓄積されますが、宿主(しゅくしゅ)たるヒトや哺乳類はそれに対応できる「HLA(=「自己」)の突然変異」を蓄積してきたのです。
その結果非常に顕著なHLA多型性になって残っています。
HLA型の表記
HLAの型は、前述したように多型に富んでいるため、血液型とは違ってやや複雑に表記されますが、この複雑さは発見の歴史に由来します。
HLAの発見後、多くの研究が世界中でなされ、それぞれの研究者はHLA抗原の多型に独自の名称を付けるようになりました。それを整理すべくWHOがスポンサーとなって命名委員会が、国際組織適合性ワークショップで規定されるようになりました。
発足当初はHLA遺伝子領域が”LA”と”Four”シリーズという異なった遺伝子から構成されているとしていたものが後にHLA-A、HLA-Bと表現されるようになりました。
このときに、当初は1から順番に番号をつけていたものが、例えば1はAの領域、5はBの領域といったように区別されたため、A座とB座には欠番が多く存在し、A座とB座で同じ番号が付けられているものはありません。後に発見されたC座は1から番号がつけられています。
また、混合リンパ球試験(MLR)で検出される第3の遺伝子に規定される抗原がMLR遺伝子(HLA-D)と命名され、さらにその後、HLA-Dと関連した抗原としてHLA-DR(D-related)抗原が命名されました。
1984年の第9回国際組織適合性ワークショップではHLA抗原を規定する遺伝子座がさらに2つ存在することがわかり、HLA-DQ、HLA-DPと命名されました。
HLAと免疫
HLAの主な働きは、自然免疫の制御、獲得免疫におけるT細胞への抗原提示です。
免疫系は様々な非自己を排除できるように複雑に構成されており、非自己の情報を得るための自己として、HLAが関与しています。
非自己のHLA、言い換えるとT細胞などから自己として認識されないHLAからは非自己の情報を得ることができず、免疫系が働きません。それどころか、自然免疫によって排除されてしまいます。
HLAと自然免疫
外来非自己(細菌、ウイルスなどの異物)が侵入すると、まずは自然免疫が働きます。自然免疫は補体が異物と複合体を形成し、これをマクロファージや単球が貪食し、さらにマクロファージが産生するIL-12によりNK細胞が強い細胞障害活性をもちます。
HLAのひとつの働きとして、NK細胞の抑制、活性化があります。
自然免疫は自己、非自己を大まかにしか判別せず、自己を寛容するためにHLAが働いています。
HLAと獲得免疫
自然免疫が働き、しばらくすると獲得免疫反応が始まります。この反応は異物を詳細に解析してオーダーメイドの排除システムを構築し、さらには異物を記憶して、再度同じ異物が侵入してきた際には効率よく排除できるようにします。獲得免疫の主役はT細胞、B細胞であり、細胞障害活性においてT細胞はグランザイムとパーフォリンといったサイトカインで、B細胞は免疫グロブリン(=Ig)を産生して攻撃します。
HLAクラスⅠ(HLA-A,B,Cなど)は、内在の9個のアミノ酸を溝に埋め込んだような形で、キラーT細胞に提示します。感染細胞などで非自己のアミノ酸が提示されると、キラーT細胞はこういった細胞を正常な部分もろとも破壊します。
HLAクラスⅡ(HLA-DR,DQ,DPなど)は、食作用などで取り込んだ外来の15個前後のアミノ酸を溝に埋め込んだような形で、ヘルパーT細胞に提示します。
ヘルパーT細胞はそれ自身は細胞障害活性をもたず、非自己のアミノ酸を確認するとTh1サイトカイン、Th2サイトカインを放出します。Th1サイトカインはキラーT細胞の活性化を促進し、Th2サイトカインはB細胞の免疫グロブリン産生を促進します。
HLAクラスⅠが体のほぼすべての細胞に存在するのに対し、HLAクラスⅡが限られた細胞(免疫担当細胞)にしか存在しないのはその働きの違いが理由です。また、キラーT細胞やヘルパーT細胞は、自己のHLAを認識して、異物の断片(アミノ酸)の情報を得ています。
